4. Autonomní diferenciální rovnice#
Co se dozvíte v tomto textu

V tomto textu se budeme věnovat diferenciálním rovnicím, kde rychlost vývoje sledované veličiny nezávisí explicitně na čase, ale jenom na veličině samotné a na parametrech prostředí. Naprostá většina modelů splňuje tyto požadavky. Důsledkem nezávislosti na čase je jistá jednoduchost při kvalitativním popisu chování řešení.
Důležitým reprezentantem autonomních diferenciálních rovnic je logistická rovnice, která bývá výchozím kamenem při studiu rostlinných a živočišných populací a označení parametrů z této rovnice písmeny \(r\) a \(K\) dalo dokonce název základním životním strategiím živočichů i rostlin.
Foto: Albatros (Diomedea exulans) v letu. Tento pták je představitelem K-stratégů. Autor JJ Harrison , https://wikimedia.org.
Show code cell source
import numpy as np
import matplotlib.pyplot as plt
import pandas as pd
from scipy.integrate import solve_ivp
Diferenciální rovnice
Rovnici je možné řešit analytickou cestou. My se nebudeme zaměřovat na hledání analytického tvaru obecného řešení, ale pokusíme se popsat chování řešení, aniž bychom tato řešení znali. Pokusíme se s co nejmenší námahou říct, jak se budou řešení chovat.
Je-li \(f(x_0)=0\), je konstantní funkce \(x(t)=x_0\) řešením rovnice. Protože derivace konstantní funkce je nula, vidíme, že řešením rovnice
\[f(x)=0\]obdržíme všechna konstantní řešení rovnice. Tato konstantní řešení se nazývají stacionární body.Stacionární body a jim odpovídající konstantní řešení představují rovnovážný stav. Často nás zajímá, jestli při vychýlení z tohoto rovnovážného stavu má systém tendenci se vrátit do původního stavu, nebo se od původního stavu dále odchylovat.
Pokud se při malém vychýlení z rovnovážného stavu systém do tohoto stavu vrací, mluvíme o stabilním stacionárním bodu.
Pokud se systém po malé výchylce do tohoto rovnovážného stavu nevrací, ale vyvíjí se k dalšímu stacionárnímu bodu nebo neohraničeně, mluvíme o nestabilním stacionárním bodu.
Následující věta umožní odlišit stabilní a nestabilní stacionární body. Protože v přírodě dochází k drobným perturbacím neustále, udává vlastně, které stacionární stavy jsou realizovatelné a můžeme je v přírodě pozorovat a které jsou prakticky nerealizovatelné.
Věta (Stabilita konstantních řešení)
Jestliže platí \(f(x_0)=0\), je konstantní funkce \(x(t)=x_0\) konstantním řešením rovnice
Protože funkce \(f\) má ve stacionárním bodě nulovou funkční hodnotu, můžeme růst nebo pokles této funkce odhalit i ze znaménka. Pokud se znaménko funkce ve stacionárním bodě mění z kladného na záporné, potom funkce klesá. V opačném případě roste.
Pravá strana bývá často rozdílem dvou faktorů, které působí proti sobě v tom smyslu, že jeden definuje rychlost růstu a druhý rychlost poklesu sledované veličiny. V takovém případě se hodí následující důsledek předchozí věty.
Důsledek (Stabilita konstantních řešení rovnice s rozdílem na pravé straně)
Jestliže platí \(f(x_0)=g(x_0)\), je konstantní funkce \(x(t)=x_0\) konstantním řešením rovnice
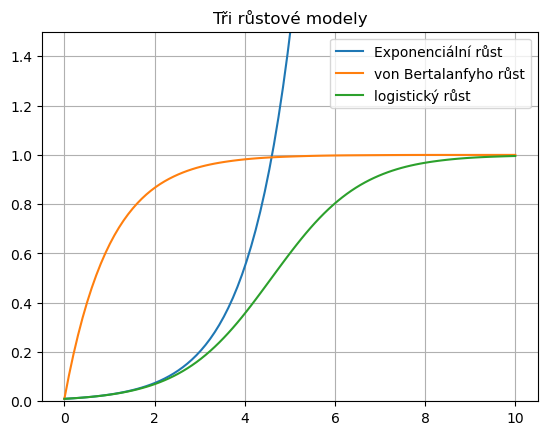
4.1. Exponenciální růst#
Nejjednodušším modelem růstu populace je model vyjadřující situaci, kdy rychlost růstu je úměrná velikosti populace. Takový růst je popsán diferenciální rovnicí
4.2. Von Bertalanffyho růst#
Dalším modelem růstu je model, kdy je růst omezen a rychlost růstu je úměrná vzdálenosti vzdálenosti od horní hranice. Původně byl formulován pro růst jedinců, kteří se mohou dožívat maximální výšky \(L_\infty\). Podle tohoto modelu je výška \(L\) dána vztahem
Kromě jedinců rostoucích rychlostí úměrnou množství chybějícímu do maximální velikosti tato rovnice popisuje i procesy, kdy změnu sledované veličiny způsobují dva faktory: růst konstantní rychlostí a pokles rychlostí úměrnou velikosti. Potom má totiž rovnice tvar
4.3. Logistický růst#
Modelem pro růst populací v prostředí s nosnou kapacitou prostředí, který je všeobecně přijímán jako vhodný kompromis mezi přesností popisu a matematickou jednoduchostí je Verhulst-Pearlův model růstu, vycházející z předpokladu, že rychlost růstu populace v prostředí s omezenou nosnou kapacitou je úměrná velikosti této populace a volnému místu v životním prostředí, nejčastěji vyjádřenému procentem z nosné kapacity. Pokud nosná kapacita prostředí je \(K\) a specifická rychlost růstu bez započtení konkurence \(r\), má model tvar
Rovnici je možno přepsat do tvaru
Poznámka (r-stratégové a K-stratégové)
Prostředí, ve kterém se populace nacházejí, je stále vystaveno změnám a výkyvům. To se odráží i na velikosti populace. Různé druhy živočichů se s těmito perturbacemi vyrovnávají různým způsobem. Některé druhy mají takřka stabilní stavy. Například populace albatrosů (Diomedea exulans) na ostrově Gough Island zůstávala téměř konstantní cca 4000 jedinců od roku 1889 přinejmenším do roku 1971 (viz [1] kapitola 5.3). Takové živočišné druhy nazýváme K-stratégy. Živočišné druhy, jejichž strategií je naopak se v příhodné době rychle rozmnožit, nazýváme r-stratégy a při modelování vývoje populace se tento faktor projevuje vysokým koeficientem \(r\). Terminologie r-strategie a K-strategie je připisována MacArthurovi a Wilsonovi, autorům teorie ostrovní biogeografie.
Více o logistické rovnici viz například [3] kapitola 6.8, [15] kapitola 1.1 a [16] str. 182.
Show code cell source
def exp_rust(t,x,k=1):
return k*x
def log_rust(t,x,r=1,K=1):
return r*x*(1-x/K)
def vB_rust(t,x,r=1,Lmax=1):
return r*(Lmax-x)
meze = [0,10]
pocatecni_podminka = [0.01]
rovnice = [exp_rust, vB_rust, log_rust]
t = np.linspace(*meze,100)
popisky = ["Exponenciální růst","von Bertalanfyho růst","logistický růst"]
data = {
popisek:
solve_ivp(
r,
meze,
pocatecni_podminka,
t_eval=t).y[0]
for popisek,r in zip(popisky,rovnice)}
df = pd.DataFrame(index=t, data = data)
ax = df.plot()
ax.set(
ylim=(0,1.5),
title="Tři růstové modely"
)
ax.grid()
ax.legend();
4.4. Autoregulace a rychlost syntézy proteinů (volitelně)#
Tento text je zpracován podle knihy [2] a pojednává o tom, jak zpětná vazba může ovlivnit produkci proteinu. Ukazuje několik strategií, jak může příroda zajistit, že v případě potřeby je protein syntetizován rychle a v dostatečném množství. Zaměříme se na matematickou stránku věci. Pěkná přednáška autora knihy a jednoho ze klíčových vědců v systémové biologii, moderní vědě, která se touto problematikou zajímá, je k dispozici na Youtube zde.
Uvažujme model syntézy proteinu, který degraduje rychlostí \(s(x)=\beta x,\) kde \(\beta\) je konstanta, tj. rychlostí úměrnou množství proteinu \(x\). To je realistický a měřením potvrzený předpoklad. Kromě toho je protein ještě i syntetizován jiným procesem. Při dosažení rovnováhy mezi degradací a produkcí je jeho úroveň konstantní.
Schopnost řízení syntézy správných proteinů ve správnou dobu je klíčová pro všechny organismy. Pro některé jednodušší organismy je vzájemný řetězec na sebe navazujících reakcí do značné míry zmapován a popsán. Tato spleť reakcí je výsledkem dlouhodobé evoluce, kde náhodné genové mutace změnily chod dosud zavedených věcí. Tyto mutace buď přinesly svému nositeli výhodu a udržely se a rozšířily, nebo naopak přinesly nevýhodu a jejich nositel mutaci dále nešířil, protože nebyl tak úspěšný v reprodukci.
V této podkapitole si ukážeme, jak se dá různými strategiemi ovlivnit rychlost, s jakou je v buňce syntetizován protein. Nejedná se o jedinou záležitost, kterou v souvislosti s chemickými reakcemi v buňce studujeme matematickými prostředky. Dalšími problémy jsou například stabilita a robustnost stacionárních stavů. Přístupy jsou jak kvalitativní (může to takto vůbec fungovat?), tak kvantitativní (dostáváme při předpovědích vypočtených z modelů správné numerické hodnoty pro pozorované veličiny?) a v obou přístupech je dovednost modelování pomocí diferenciálních rovnic nezastupitelnou pomůckou.
4.4.1. Produkce konstantní rychlostí#
Množství proteinu označme \(x\) a předpokládejme produkci konstantní rychlostí. Spolu s degradací z předchozího odstavce tedy máme model
4.4.1.1. Nondimenzionalizace#
Vhodnou volbou jednotek pro veličinu \(x\) je možné docílit toho, že platí \(\alpha = \beta.\) Opravdu, rovnici je možno přepsat na tvar
4.4.1.2. Růstové křivky#
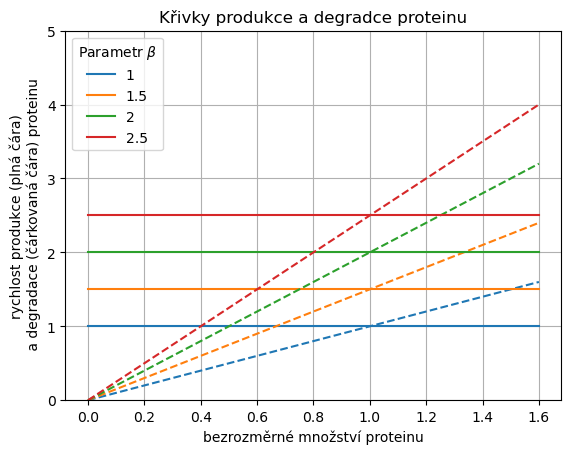
Nejprve nakreslíme křivky produkce a degradace do jednoho obrázku, abychom dokázali lokalizovat stacionární body.
Křivky pro stejné hodnoty parametru jsou stejnou barvou.
Průsečík křivek stejné barvy je stacionární bod systému při nastavení parametrů daném příslušnou barvou. Všimněte si, že všechny tyto průsečíky jsou nad sebou a každý v jiné výšce. Pro všechny parametry nastává stacionární stav pro stejné množství proteinu \(x=1\), ale rychlosti produkce a degradace se v tomto stacionárním stavu liší.
Všechna nastavení parametrů vykazují převažující růst pro malé a převažující degradaci pro velké hodnoty \(x\). Stacionární body jsou tedy stabilní.
Show code cell source
beta = [1,1.5,2,2.5] # definice parametrů
x = np.linspace(0, 1.6, 100) # definice intervalu pro kreslení růstových křivek
fig, ax = plt.subplots()
for i,beta_ in enumerate(beta): # Cyklus přes parametry
produkce = beta_+0*x # Křivka produkce
degradace = beta_*x # Křivka degradace
ax.plot(x,produkce, color="C"+str(i), label=beta_) # Vykreslení křivky produkce
ax.plot(x,degradace, "--", color="C"+str(i), label=None) # Vykreslení křivky degradace
ax.set(title="Křivky produkce a degradce proteinu",
xlabel='bezrozměrné množství proteinu',
ylabel='rychlost produkce (plná čára)\n a degradace (čárkovaná čára) proteinu',
ylim=(0, 5)
);
plt.legend(
title=r"Parametr $\beta$"
)
plt.grid();
4.4.1.3. Časový vývoj řešení#
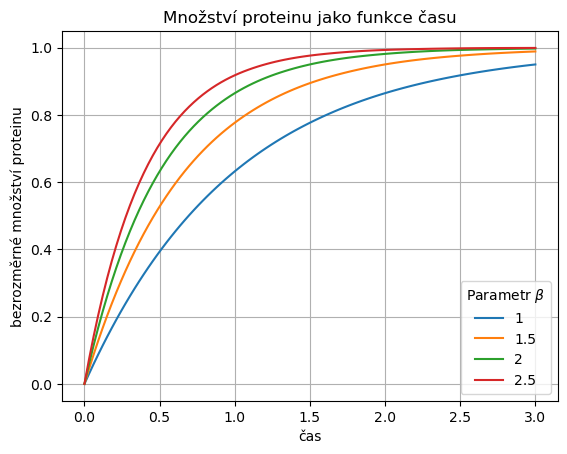
Průběh řešení pro jednotlivé hodnoty parametru u řešení vycházejícího z nulové počáteční podmínky je na obrázku. V praxi nás zajímá, jak rychle bude dosaženo určitého množství proteinu. Tedy vybereme si nějakou hladinu, například \(0.6\), a sledujeme, která křivka této hladiny dosáhne jako první a která jako poslední.
Show code cell source
meze = [0,3] # definice počátečního a koncového časového okamžiku
t = np.linspace(*meze, 100) # definice intervalu pro kreslení funkcí času
reseni = np.array([
solve_ivp(
lambda t, x: beta_ * (1 - x),
meze,
[0],
t_eval=t).y[0]
for beta_ in beta
]).T
plt.plot(t,reseni)
ax = plt.gca()
ax.set(
title="Množství proteinu jako funkce času",
xlabel='čas',
ylabel='bezrozměrné množství proteinu',
)
plt.grid()
plt.legend(
beta,
title=r"Parametr $\beta$"
);
4.4.1.4. Rychlá produkce je „zaplacena“ rychlou degradací#
Je patrné, že nejrychleji hladina enzymu roste pro nejvyšší hodnotu parametru. Vodorovnou čáru ve výšce \(0.6\) protne jako první křivka odpovídající nejvyšší hodnotě parametru \(\beta\). Být první je neskutečně důležité, někdy přímo otázka života a smrti. Taková bakterie, která umí rychle produkovat enzym, například umí rychle uniknout z prostředí s nepříznivými životními podmínkami (rychle sestaví proteiny, které vytvoří jakýsi molekulární motor a ten buňku posune na jiné místo) nebo umí rychle začít ukládat v přítomnosti glukózy energii pro budoucí využití. Na druhou stranu, popsaným mechanismem je rychlost je dosažena nejenom rychlou produkcí, ale i rychlou degradací proteinu. Tedy protein se rychle produkuje a rychle rozpadá. Už z laického hlediska se takový postup nezdá být postupem optimálním pro přežití. Je to jako bychom u auta chtěli mít možnost okamžitě zrychlit a za tímto účelem udržovali vysoké otáčky motoru, ale abychom jeli rozumnou rychlostí tak bychom brzdili. Tedy bychom současně šlapali na plyn i na brzdu. To rozumný člověk nedělá.
Proteiny ve skutečnosti nejsou rychle odbourávány. Popsaný mechanismus šlapání na brzdu a plyn současně v buněčném světě ve skutečnosti není použit. Životnost proteinu není řádově odlišná od životnosti buňky. Příroda a evoluce by takové plýtvání nepřipustily a namísto toho se k problému rychlé produkce proteinu staví jinak. Použitím zpětné vazby, což je předmětem následující kapitoly.
4.4.1.5. Bezrozměrná formulace#
Zkusíme ještě přepsat rovnici s konstantní produkcí do bezrozměrných jednotek. Rovnici
Show code cell source
for i,beta_ in enumerate(beta):
plt.plot(t*beta_,reseni[:,i],label=beta_,lw=(10-2*i),alpha=0.3)
ax = plt.gca()
ax.set(
xlabel="bezrozměrný čas",
ylabel="bezrozměrné množství proteinu",
title="Syntéza proteinu v bezrozměrných jednotkách"
)
plt.legend();
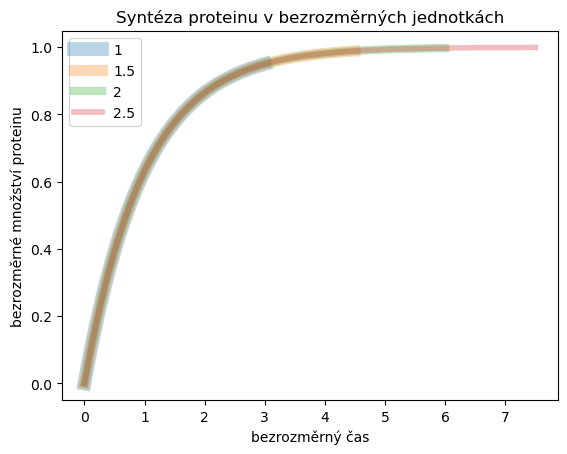
Vidíme názorně výhodu bezrozměrných jednotek. Není nutné řešit rovnici pro různé hodnoty parametru, ale všechna řešení jsou až na transformaci stejná. Jinými slovy vyjádřeno, v modelu
4.4.2. Produkce se zápornou zpětnou vazbou#
Předpokládejme produkci proteinu rychlostí související s množstvím proteinu. Jedná se tedy o model
Vhodný tvar funkce \(f(x)\) může být například rozdíl konstanty a třetí mocniny, tj.
Čím vyšší hodnota koeficientu \(\alpha\), tím je počáteční produkce rychlejší a dříve je dosaženo potřebného množství enzymu. Toto se děje při konstantní zpětné vazbě a konstantním stacionárním stavu (všechny rovnice mají člen popisující degradaci ve tvaru \(-x\) a stacionární stav \(x=1\)).
4.4.2.1. Nondimenzionalizace#
Vhodnou volbou jednotek pro veličinu \(x\) dosáhneme toho, že stacionárním bodem bude \(x=1\) (jinými slovy, množství enzymu budeme měřit v procentech rovnovážného stavu). Protože budeme uvažovat stále stejnou rychlost degradace a sledovat jenom vliv funkce \(f(x)\), můžeme jednotky času zvolit tak, že \(\beta = 1\). Tj. studujeme rovnici
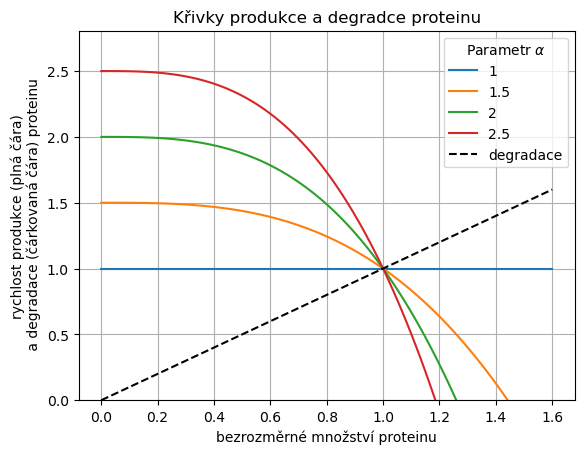
4.4.2.2. Křivky růstu a degradace#
Funkce
Show code cell source
alpha = [1,1.5,2,2.5] # definice parametrů
x = np.linspace(0, 1.6, 100) # definice intervalu pro kreslení růstových křivek
fig, ax = plt.subplots()
for i,alpha_ in enumerate(alpha): # Cyklus přes parametry
produkce = alpha_ - (alpha_-1)*x**3 # Křivka produkce
ax.plot(x,produkce, color="C"+str(i), label=alpha_) # Vykreslení křivky produkce
ax.plot(x,x, "--", color="k", label="degradace") # Vykreslení křivky degradace
ax.set(title="Křivky produkce a degradce proteinu",
xlabel='bezrozměrné množství proteinu',
ylabel='rychlost produkce (plná čára)\n a degradace (čárkovaná čára) proteinu',
ylim=(0, None)
);
plt.legend(
title=r"Parametr $\alpha$"
)
plt.grid();
4.4.2.3. Časový průběh řešení modelu#
Zkusíme si namodelovat průběh řešení modelu pro různé hodnoty parametru.
Show code cell source
meze = [0,3] # definice počátečního a koncového časového okamžiku
t = np.linspace(*meze, 100) # definice intervalu pro kreslení funkcí času
reseni = np.array([
solve_ivp(
lambda t, x: alpha_ - (alpha_-1)*x**3 - x,
meze,
[0],
t_eval=t).y[0]
for alpha_ in alpha
]).T
plt.plot(t,reseni)
ax = plt.gca()
ax.set(
title="Množství proteinu jako funkce času",
xlabel='čas',
ylabel='bezrozměrné množství proteinu',
)
plt.grid()
plt.legend(
alpha,
title=r"Parametr $\alpha$"
);
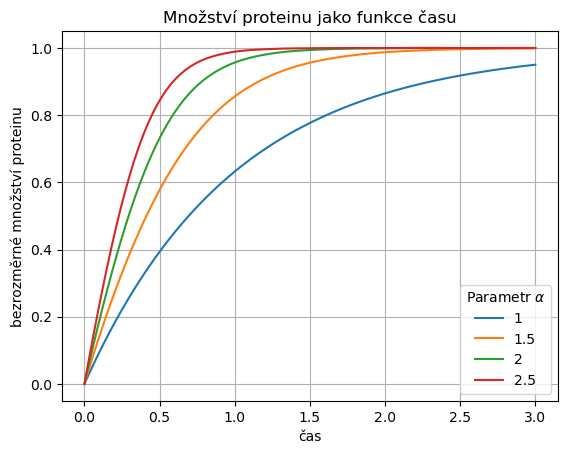
Čím vyšší hodnota koeficientu \(\alpha\), tím je počáteční produkce rychlejší a dříve je dosaženo potřebného množství enzymu. Toto se děje při konstantní zpětné vazbě a konstantním stacionárním stavu (všechny rovnice mají člen popisující degradaci ve tvaru \(-x\) a stacionární stav \(x=1\)).
4.4.2.4. Negativní zpětná vazba urychluje syntézu#
Vidíme, že negativní zpětná vazba příznivě ovlivňuje rychlost syntézy proteinu. Tím je buňce umožněno zareagovat rychle na vnější podmínky, aniž by za to platila cenou rychlé degradace, jako bez zpětné vazby (současně přidávání plynu a brzdění). Taková zpětná vazba může být zprostředkována například tím, že protein vstupuje do jiných chemických reakcí.
Příroda zařadila i do jednoduchých organismů typu buňka takových zpětných vazeb obrovské množství a dlouholetou evolucí vyladila hodnoty parametrů tak, aby potřebné proteiny vznikaly přesně v době, kdy jsou potřeba a aby dostatečně rychle vznikly v potřebné koncentraci. To je zařízeno mutacemi v genech. Chybou v přepise genů například může vzniknout zpětná vazba, která do té doby neexistovala. Pokud se projeví jako užitečná, dává svému nositeli evoluční výhodu a šíří se dál. Podobně se během milionů let vyladí parametry této zpětné vazby, například hodnota parametru \(\alpha\). Těmito problémy zabývá věda nazývaná systémová biologie. Tato věda prodělala bouřlivý rozvoj v letech okolo roku 2000, kdy byly vyvinuty účinné postupy pro získávání potřebných dat o genech a proteinech a tento rozvoj pokračuje dodnes.
Poznatky, na kterých je založen tento model, jsou potvrzeny experimentálně. Používá se například vnesení genu způsobujícího světélkování do genetické informace bakterií.